It’s halfway through the month of July and I realized I haven’t updated this blog since April! There are some things I can talk about in more detail in the future, but for now, I will just give a basic summary of my life updates since the last time I posted here.
First, this spring quarter, the last quarter of my first year of graduate school, was challenging but rewarding in a number of ways. I made some significant progress on my research project, and I feel that I gained a lot of valuable experience in conducting independent research, programming, and data presentation. Although I had a full course load, I tried to set aside enough time to work on research each week so that I would have something to present at our weekly group meetings, and my PI seemed impressed with my progress. He gave me some suggestions for developing what I have so far, but said that during the summer I could think about starting a paper draft, which is pretty exciting. I have been spending a lot more time in the lab and getting to know the other grad students and postdocs who work there, and I feel a sense of community with them much more so than what I felt in my undergrad research group. My course load this past quarter consisted of two neuroscience courses and one biomedical data analysis course. The neuroscience courses were more challenging than the systems neuroscience course I took in winter quarter, but were extremely valuable in providing me with more of the biological background as well as a deeper understanding of how experiments work. I also made valuable connections with my classmates and some of the guest lecturers. I have been continuing conservations with two professors I met through these courses, as there might be a potential for setting up collaborations.
At the end of next month, August, I will have a series of five qualifying examinations, of which I must pass at least 3 in order to continue in the program. These exams are only offered once a year, and students get two tries to pass them. Ideally, I will be able to pass them in the first round, this August, because it will be a huge weight lifted off my shoulders, and that way in my second year, I can fully focus on my new elective courses and research rather than worrying about reviewing the material from the core courses. For the first half of the summer so far, I spent part of the time reviewing all my notes in these five subjects, and part of the time working on image analysis in the lab and connecting with neuroscience professors. My plan is that after today, I will completely direct my focus to the qualifying exams. After the last set of exams end in August, I will have almost a full month before courses start for the fall. During that break period, I can work in the lab full time and start thinking about writing a paper draft.
This first half of the summer has been both enjoyable and meaningful and has helped me organize my goals and remind myself of the things that are important to me. Since classes ended, I have been going into the lab during weekdays from 10 am to 5 pm, spending half the time on research and half the time on studying for quals. In the evenings, I had time to go to the gym, run outside, and swim, grab dinner and catch up with friends from college who are around during the summer, hang out with my roommate, play with my dog, and work on some painting. I felt balanced and in control of my schedule, and it reminded me of how much I’m looking forward to when my classes are over and I can fully focus on research during the day and hopefully get a little downtime in the evening. I feel lucky for the fact that both my PI and the postdoc who works in my subgroup seem to be advocates for work-life balance. In undergrad, I spent most of my time studying and working on research, and I didn’t feel like I was taking care of my physical and mental health as much, but coming to grad school, I have come to appreciate the value of balance, both for short-term work efficiency and long-term health. In the next month and a half, I will be focusing a lot on the qualifying exams, and although I hope to continue my workout schedule and play with my dog a little in the evenings, I will probably cut down a lot of socializing and painting during this period. I feel motivated by thinking about how good I’ll feel after passing and how nice my life could be after getting through this initial hurdle.
The past couple of months, I have been a lot more social than I usually am. It is interesting because recently, a lot of people who are important to me have come out to me as members of the LGBTQ+ community, possibly because I have been more out in social spaces and become more comfortable embracing my own identity. I have realized that there are a lot more queer people than I originally thought, and it feels a lot less isolating knowing that I can relate to people in ways that I didn’t know before. One of my friends came to visit me during the weekend when one of the queer orgs I am a part was having a social event in a drag cafe in West Hollywood, and we had a great time watching the show and going bar hopping later. I think I enjoyed the experience a lot more than I usually enjoy those settings because she is someone that I feel very comfortable with and can connect with on a lot of other levels outside just queerness. This past weekend, she and one of my other friends who recently came out to me joined me on a road trip to San Diego for the Pride Parade and Festival. When we were there, we met up with a friend I met through my undergrad research group, and the three of them got along with each other surprisingly well. San Diego Pride is special to me because so much of my coming out process began there, and it was in that city I feel I truly began my growth as a person rather than just as a student. In addition to the parade in Hillcrest and the festival in Balboa Park, I was able to visit a lot of the old spots in San Diego that I enjoy, such as Chocolat, a gelato shop in Gaslamp, Downtown San Diego, The Living Room, a hookah lounge and restaurant in Downtown La Jolla where I spent many days and nights studying during undergrad, and Gossip Grill, the only girl bar I know of. One of my favorite queer female artists, King Princess, was performing at the festival, and it was incredible to watch her live and sing along to the songs that I had heard so many times in my own car.
Another event that I recently participated in this summer was the LGBTQ+ STEM Day, a conference in the LA LGBT Center that connected a lot of universities in California. I found out about it through QSTEM, one of the orgs I participated in, and I challenged myself to participate in the open mic, where people could give five-minute research talks. It was my first time speaking about my research in a setting where most people are not from my field, but I think I was able to connect with the audience. After the talks, an undergrad I vaguely know from QSTEM came to talk to me about his research interests and how they relate to my project. I told him about the PI I worked for when I was an undergrad, since his interests seemed to align well with the focus of that group, and he is considering graduate school sometime in the future. I also ran into a woman I had met three years ago when I was taking a summer class in undergrad. She was taking classes in San Diego during that summer, but she is currently a student in my grad school institution in Los Angeles. She introduced me to some of her friends there, and a few of us ended up hanging out during the fourth of July. Overall, it was a meaningful experience to connect to fellow queer people in STEM and challenge myself to speak publicly about my research. I hope to look into more opportunities to present my research and network in the future, such as conferences and other LGBTQ+ STEM events.
Today, I deleted all my social media apps and am planning to limit my screen time until after qualifying exams, as I would really like to get through them during this round. I hope that I can find ways to stay motivated and enjoy the process of learning this material, as a familiarity with the vast tools in applied mathematics will probably serve me not only for these exams, but for my research progress in the future. During the pride roadtrip, an interesting point one of my friends said was that success is often a result of the combination of three main factors: 1) prioritization of long-term goals over short-term satisfaction, 2) insecurity about ones’ current position and a desire to improve, and 3) a slight sense of elitism. I thought this was extremely interesting and somewhat helpful in guiding the remainder of my quals studying process. Deleting social media apps and limiting hang out sessions and painting are examples of prioritizing my long term goal of passing the quals over short-term pleasure, and the fact that I am not sure whether or not I will pass and be able to continue in the program is an example of the insecurity piece. I think the elitism piece is also interesting, and I definitely believe that some level of overconfidence in one’s natural abilities can be helpful for performance. When I was in undergrad, my grad student mentor once told me about this exercise she saw where women in STEM were asked to write down good things about themselves before taking exams, and it actually helped boost their performance. I tried this a few times and it actually did help improve my headspace before exams a lot. Thus, I think that it will be helpful for me to keep a notebook during my quals studying process, and every now and then try to write down things that remind me of my own strengths, but not quite to the extent that I no longer have the anxiety about passing and the motivation to study harder.
That is all I have for this post – I would post group photos from queer events, but I would like to respect the privacy of the other people in the photos. Hopefully after the exams in August, I can update more frequently and maybe make more science posts about my research process. Until then, I really hope I can get through these quals!




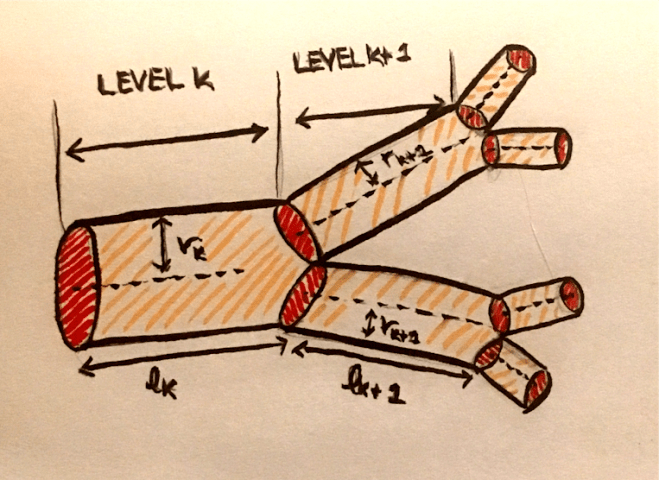

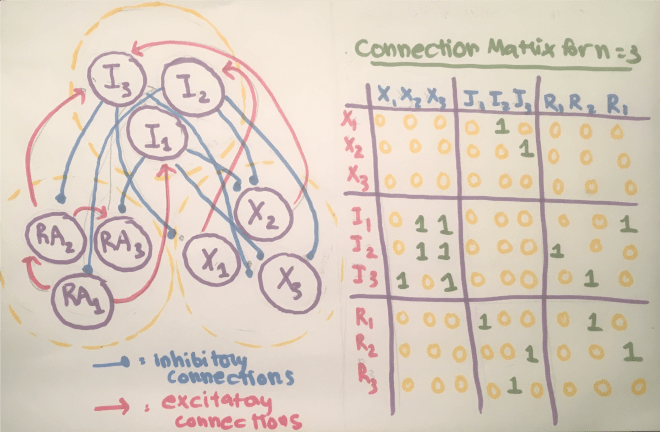
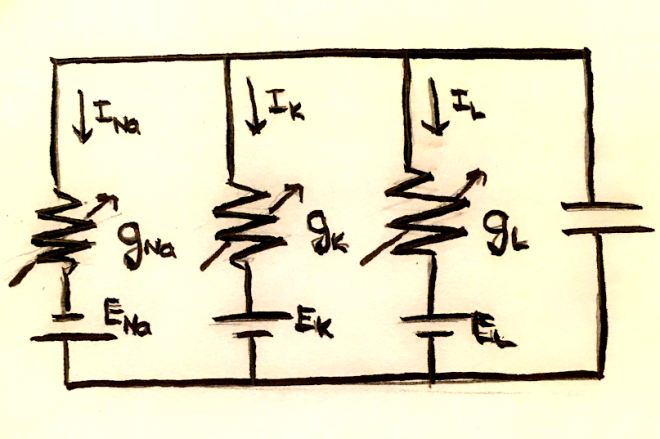 Because of charge conservation, the sum of the currents across the capacitor and each of the resistors must be 0. In mathematical terms, this is $C_m \frac{dV_m}{dt} = -\sum_i I_i$.
Because of charge conservation, the sum of the currents across the capacitor and each of the resistors must be 0. In mathematical terms, this is $C_m \frac{dV_m}{dt} = -\sum_i I_i$.